Gibourtia copallifera is a rare, range-restricted tree native to the Kasewe Forest Reserve in southern Sierra Leone. Historically exploited for gum copal, it now faces threats from charcoal production due to its high-quality charcoal. Although the species shows good growth in forest patches established 60 years ago, natural regeneration remains minimal. While many tropical tree species form beneficial relationships with arbuscular mycorrhizal fungi (AMF), these associations are still poorly understood in humid regions like Sierra Leone. This study investigated the AMF associations of G. copallifera and assessed changes in mycorrhizal diversity in relation to forest degradation and tree development stages. Soil characterization was conducted through profile analysis, and samples were subjected to laboratory testing. Three soil types were identified within the reserve: K 01—barren land with sparse vegetation; K 02—forest land dominated by G. copallifera; and K 03—upland fallow agricultural land with shrubs and a few trees. Soil analysis indicates pH values ranging from 5.03 to 5.87 (acidic), with calcium as the most dominant exchangeable base, followed by magnesium, potassium, and sodium. The surface horizon under G. copallifera exhibited the highest total exchangeable bases, and high cation exchange capacity (CEC) was linked to the presence of decomposed plant matter. A total of 22 AMF species were identified in plant root samples, with a significantly higher proportion found in non-degraded forest patches. These accounted for around 68% of AMF species, especially Scutellospora and Gigaspora, along with Glomus and Acaulospora. AMF species richness and diversity were considerably higher in non-degraded patches (3.13 species) than in degraded areas (1.75). Mycorrhizal frequency and intensity were also significantly greater in undisturbed forest sites. AMF colonization peaked in mature trees and was lowest in seedlings, although the variation across trials was not statistically significant (P = 0.07). The study concludes that K01 is unsuitable for G. copallifera due to its shallow depth, which restricts growth. In contrast, K02 and K03 are more favorable for the species. However, many K02 areas have been converted to agricultural land, and in locations where natural forest regeneration is allowed, more aggressive species such as Gmelina arborea and Anisophyllea laurina tend to dominate. Additionally, AMF play a crucial role in promoting the growth of G. copallifera in nutrient-poor soils. These findings are important for informing reintroduction and reforestation strategies for this native tropical tree species.
| Published in | Plant (Volume 13, Issue 3) |
| DOI | 10.11648/j.plant.20251303.12 |
| Page(s) | 124-137 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Mycorrhizal, Arbuscular, Forest, Soil Fertility, Guibourtia, Degraded, Reserve, Sierra Leone
Property | Type 1 | Type 2 | Type 3 | |||
|---|---|---|---|---|---|---|
Soil Depth cm | 0-10 | 0-10 | 10 to 70 | 0-10 | 10 to 50 | 50-100 |
Gravel | 92 | 3.8 | 92 | 75 | 89 | 91 |
Sand | 88 | 80 | 78 | 82 | 84 | 86 |
Silt | 4 | 8 | 8 | 8 | 6 | 6 |
Clay | 8 | 12 | 14 | 10 | 10 | 8 |
pH | 5.87 | 5.82 | 5.09 | 5.69 | 5.03 | 5.77 |
EC | 58.5 | 56.9 | 25.1 | 64.2 | 19.1 | 98.9 |
C | 4.52 | 5.52 | 2.84 | 4.52 | 2.08 | 2.08 |
N | 0.01 | 0.08 | 0.03 | 0.04 | 0.01 | 0.01 |
P mg/kg soil | 9.48 | 4.82 | 8.73 | 8.66 | 8.81 | 7 |
Na mg/kg soil | 2.3 | 2.8 | 2.1 | 2.5 | 2.1 | 2.8 |
K mg/kg soil | 4.9 | 6.9 | 3 | 5.92 | 1.65 | 2.01 |
Mg mg/kg soil | 16.6 | 18.6 | 10.1 | 12.1 | 14.6 | 13.4 |
Ca mg/kg soil | 26 | 28.2 | 10.1 | 26.54 | 21.41 | 14.22 |
CEC cmol(+)/kg soil | 10.56 | 12.86 | 10 | 11.24 | 8.07 | 7.28 |
K 01 | K 02 | K 03 | |
|---|---|---|---|
0 to 10 cm | Very dark yellowish brown (10YR3/2), gravelly loamy sand, weak, coarse, granular structure and friable, non-sticky, non-plastic consistence. Many pores and roots | Reddish brown (10YR3/2), sandy loam, weak, coarse, sub-angular blocky structure and friable, non-sticky, non-plastic consistence. Many fine pores and few coarse pores many roots | Yellowish brown (10YR3/2), gravelly sandy clay loam, moderate, medium, sub-angular blocky structure and friable, slightly sticky, slightly plastic consistence |
10 to 70 cm | Impenetrable | Yellowish brown (10YR4/6), gravelly sandy loam, moderate, fine, sub-angular blocky structure and friable, slightly sticky, slightly plastic. Many pores, few roots | Yellowish brown (10YR4/6), gravelly sandy clay, weak, medium, sub-angular blocky structure and friable, sticky, plastic consistence |
70 to 100 cm | Impenetrable | Impenetrable | Dark yellowish brown, gravelly Sandy clay, weak, medium, sub-angular blocky structure and friable, sticky, plastic consistence |
Family/Genera | Non-Degraded Patch | Degraded | No. of Species |
|---|---|---|---|
Glomeraceae Glomus spp | 8 | 5 | 10 |
Acaulosporaceae Acaulospora spp | 6 | 3 | 7 |
Acaulosporaceae Kuklospora spp | 1 | 2 | 2 |
Gigasporaceae Gigaspora spp | 2 | 1 | 2 |
Gigasporaceae Scutellospora spp | 4 | 2 | 3 |
Entrophosporaceae Entrophospora spp | 1 | 0 | 1 |
Ambisporaceae Ambispora spp | 2 | 1 | 2 |
Paraglomeraceae Paraglomus spp | 1 | 0 | 1 |
totals | 25 | 14 | 28 |
SUMMARY | Count | Sum | Average | Variance |
|---|---|---|---|---|
Seedling | 10 | 687 | 68.7 | 18.45556 |
Sapling | 10 | 2218 | 221.8 | 1642.844 |
Juvenile | 10 | 1642 | 164.2 | 3501.067 |
Young Tree | 10 | 3832 | 383.2 | 2941.289 |
Matured Tree | 10 | 4340 | 434 | 8189.111 |
Day_1 | 5 | 1497 | 299.4 | 31148.3 |
Day_2 | 5 | 1424 | 284.8 | 44640.2 |
Day_3 | 5 | 1315 | 263 | 22446 |
Day_4 | 5 | 1263 | 252.6 | 21824.3 |
Day_5 | 5 | 1118 | 223.6 | 18474.3 |
Day_6 | 5 | 1219 | 243.8 | 21711.7 |
Day_7 | 5 | 1385 | 277 | 29520.5 |
Day_8 | 5 | 1173 | 234.6 | 23109.3 |
Day_9 | 5 | 1278 | 255.6 | 31590.3 |
Day_10 | 5 | 1047 | 209.4 | 14745.3 |
SUMMARY | Count | Sum | Average | Variance |
|---|---|---|---|---|
T_1 | 5 | 475 | 95 | 3605.5 |
T_2 | 5 | 483 | 96.6 | 2973.8 |
T_3 | 5 | 518 | 103.6 | 3584.8 |
T_4 | 5 | 506 | 101.2 | 3562.7 |
T_5 | 5 | 386 | 77.2 | 2466.7 |
T_6 | 5 | 386 | 77.2 | 2224.2 |
T_7 | 5 | 446 | 89.2 | 2829.7 |
T_8 | 5 | 390 | 78 | 2698 |
T_9 | 5 | 362 | 72.4 | 2041.3 |
T_10 | 5 | 418 | 83.6 | 2298.8 |
T_11 | 5 | 450 | 90 | 2655.5 |
T_12 | 5 | 351 | 70.2 | 2008.7 |
T_13 | 5 | 430 | 86 | 2606 |
T_14 | 5 | 374 | 74.8 | 2607.2 |
T_15 | 5 | 429 | 85.8 | 2429.2 |
T_16 | 5 | 389 | 77.8 | 2265.7 |
T_17 | 5 | 458 | 91.6 | 2617.3 |
T_18 | 5 | 405 | 81 | 2563.5 |
T_19 | 5 | 434 | 86.8 | 2706.7 |
T_20 | 5 | 376 | 75.2 | 2862.2 |
T_21 | 5 | 432 | 86.4 | 3102.3 |
T_22 | 5 | 469 | 93.8 | 3370.7 |
T_23 | 5 | 454 | 90.8 | 3117.2 |
T_24 | 5 | 438 | 87.6 | 3006.3 |
T_25 | 5 | 415 | 83 | 3038 |
T_26 | 5 | 443 | 88.6 | 2348.8 |
T_27 | 5 | 420 | 84 | 2931.5 |
T_28 | 5 | 354 | 70.8 | 1665.2 |
T_29 | 5 | 452 | 90.4 | 2682.8 |
T_30 | 5 | 406 | 81.2 | 1992.2 |
Seedling | 30 | 689 | 23.0 | 7.0 |
Sapling | 30 | 2246 | 74.9 | 218.0 |
Juvenile | 30 | 1642 | 54.7 | 122.4 |
young tree | 30 | 3832 | 127.7 | 355.9 |
Matured tree | 30 | 4340 | 144.7 | 211.7 |
SUMMARY | Count | Sum | Avera/ge | Variance |
|---|---|---|---|---|
Seedling | 10 | 2793 | 279.3 | 12744.7 |
Sapling | 10 | 2991 | 299.1 | 8616.5 |
Juvenile | 10 | 3680 | 368.0 | 6745.1 |
Young Tree | 10 | 4969 | 496.9 | 5973.0 |
Matured Tree | 10 | 5679 | 567.9 | 18315.2 |
Day_1 | 5 | 2152 | 430.4 | 14286.8 |
Day_2 | 5 | 2298 | 459.6 | 49965.8 |
Day_3 | 5 | 1928 | 385.6 | 28887.8 |
Day_4 | 5 | 1804 | 360.8 | 47254.7 |
Day_5 | 5 | 1714 | 342.8 | 10236.7 |
Day_6 | 5 | 1834 | 366.8 | 10103.2 |
Day_7 | 5 | 1652 | 330.4 | 10868.8 |
Day_8 | 5 | 2414 | 482.8 | 28644.2 |
Day_9 | 5 | 2207 | 441.4 | 30494.8 |
Day_10 | 5 | 2109 | 421.8 | 14923.7 |
SUMMARY | Count | Sum | Average | Variance |
|---|---|---|---|---|
T_1 | 5 | 721 | 144.2 | 3686.7 |
T_2 | 5 | 665 | 133 | 1683.5 |
T_3 | 5 | 618 | 123.6 | 1846.8 |
T_4 | 5 | 677 | 135.4 | 2514.3 |
T_5 | 5 | 610 | 122 | 2224.5 |
T_6 | 5 | 693 | 138.6 | 2718.3 |
T_7 | 5 | 687 | 137.4 | 3463.3 |
T_8 | 5 | 652 | 130.4 | 3028.3 |
T_9 | 5 | 622 | 124.4 | 3268.3 |
T_10 | 5 | 678 | 135.6 | 3253.3 |
T_11 | 5 | 656 | 131.2 | 2902.7 |
T_12 | 5 | 688 | 137.6 | 1919.8 |
T_13 | 5 | 677 | 135.4 | 2087.8 |
T_14 | 5 | 687 | 137.4 | 1406.3 |
T_15 | 5 | 683 | 136.6 | 2390.3 |
T_16 | 5 | 708 | 141.6 | 1456.3 |
T_17 | 5 | 667 | 133.4 | 2558.3 |
T_18 | 5 | 718 | 143.6 | 2629.3 |
T_19 | 5 | 829 | 165.8 | 2708.2 |
T_20 | 5 | 645 | 129 | 1618.5 |
T_21 | 5 | 722 | 144.4 | 6863.3 |
T_22 | 5 | 688 | 137.6 | 2424.3 |
T_23 | 5 | 584 | 116.8 | 3358.7 |
T_24 | 5 | 675 | 135 | 2846.5 |
T_25 | 5 | 734 | 146.8 | 3150.7 |
T_26 | 5 | 734 | 146.8 | 1587.2 |
T_27 | 5 | 670 | 134 | 1702 |
T_28 | 5 | 724 | 144.8 | 2142.2 |
T_29 | 5 | 669 | 133.8 | 1776.7 |
T_30 | 5 | 689 | 137.8 | 3935.7 |
Seedling | 30 | 2335 | 77.8 | 155.5 |
Sapling | 30 | 3112 | 103.7 | 270.5 |
Juvenile | 30 | 4088 | 136.3 | 234.3 |
Young Tree | 30 | 4879 | 162.6 | 352.8 |
Matured Tree | 30 | 6056 | 201.9 | 524.2 |
AMF | Arbuscular Mycorrhizal Fungi |
CEC | Cation Exchange Capacity |
NTFPs | Non-timber Forest Products |
EC | Electrical Conductivity |
KOH | Potassium Hydroxide |
HCl | Hydrochloric Acid |
H2O2 | Hydrogen Peroxide |
RLC | Root Length Colonization |
ANOVA | Analysis of Variance |
SEB | Sum of the Exchanged Bases |
SOM | Soil Organic Matter |
K 01 | Soil Type 1 |
K 02 | Soil Type 2 |
K 03 | Soil Type 3 |
AM | Arbuscular Mycorrhizal |
SOC | Soil Organic Carbon |
| [1] | Manucci PM, Franchini M. 2017. Health effects of ambient air pollution in developing countries. Int J Environ Res Public Health. 14: 10-22. |
| [2] | Govindan, M., Rajeshkumar, P. P., Varma, C. K. Y., Anees, M. M., Rashmi, C. R., and Nair, A. B. 2020. Arbuscular mycorrhizal fungi status of mango (Mangifera indica) cultivars grown in typic quartzipsamments soil. Agric. Res. 9, 188-196. |
| [3] | Adamo, I., Castaño, C., Bonet, J. A., Colinas, C., Martı́nez de Aragón, J., and Alday, J. G. 2021. Soil physico-chemical properties have a greater effect on soil fungi than host species in Mediterranean pure and mixed pine forests. Soil Biol. Biochem. 160, 108320. |
| [4] | Sokołowska, J.; Józefowska, A.; Wóznica, K.; Zaleski, T 2020. Succession from meadow to mature forest: Impacts on soil biological, chemical and physical properties—Evidence from the Pieniny Mountains, Poland. Catena 2020, 189, 104503. |
| [5] | Crowther TW, Van den Hoogen J, Wan J, Mayes MA, Keiser A, Mo L, 2019. The global soil community and its influence on biogeochemistry. Science 2019; 365. |
| [6] | Bueno CG, Moora M, Gerz M, Davison J, Öpik M, Pärtel M, 2017. Plant mycorrhizal status, but not type, shifts with latitude and elevation in Europe. Glob Ecol Biogeo, 26: 690–9. |
| [7] | Steidinger BS, Crowther TW, Liang J, Nuland MEV, Werner GDA, Reich PB. 2019. Climatic controls of decomposition drive the global biogeography of forest-tree symbioses. Nature 2019; 569: 404–8. |
| [8] | Dixon RK, Garrett HE, Cox GS, Marx DH, Sander IL. 1984. Inoculation of three Quercus species with eleven isolates of ectomycorrhizal fungi. I. inoculation success and seedling growth relationships. Science. 1984; 30: 364–72. |
| [9] | Bennett JA, Maherali H, Reinhart KO, Lekberg Y, Hart MM, Klironomos J. 2017. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science, 355: 181–4. |
| [10] | Abalos, D., van Groenigen, J. W., Philippot, L., Lubbers, I. M., De Deyn, G. B., 2019. Plant trait-based approaches to improve nitrogen cycling in agroecosystems. Journal of Applied Ecology 56, 2454–2466. |
| [11] | Diogo IJS, Fortunato MEM, Costa IR 2015. Seed deposition in the edge-interior gradient of a degraded fragment of tropical semideciduous forest, Northeastern Brazil. Revista de Biología Tropical 63(4): 981-994. (Int. J. Trop. Biol. ISSN-0034-7744). |
| [12] | FAO. 2015. News detail. Food and Agriculture Organization of the United Nations. |
| [13] | Lozano-Baez, S. E.; Cooper, M.; Meli, P.; Ferraz, S. F. B.; Rodrigues, R. R.; Sauer, T. J. 2019. Land restoration by tree planting in the tropics and subtropics improves soil infiltration, but some critical gaps still hinder conclusive results. For. Ecol. Manag. 2019, 444, 89–95. |
| [14] | De Beenhouwer M, Muleta D, Peeters B, Van Geel M, Lievens B, Honnay O. 2015a. DNA pyrosequencing evidence for large diversity differences between natural and managed coffee mycorrhizal fungal communities. Agronomy for Sustainable Development 35: 241–249. |
| [15] | Ohsowski BM, Zaitsoff PD, Opik M, Hart MM. 2014. Where the wild things are: looking for uncultured Glomeromycota. New Phytologist 204: 171–179. |
| [16] | Bakarr and Janos 1996. Grandy, A. S., Wieder, W. R., Wickings, K., Kyker-Snowman, E., 2016. Beyond microbes: 716 Are fauna the next frontier in soil biogeochemical models? Soil Biology and 717 Biochemistry, Special issue: Food web interactions in the root zone: influences on community and ecosystem dynamics 102, 40–44. |
| [17] | Munro PG, van der Horst GA 2012a. The Domestic Trade in Timber and Fuelwood Products in Sierra Leone: Current Dynamics and Issues. Freetown: FAO/EU. |
| [18] | Munro P, van der Horst G, Healy S. 2017. Energy justice for all? Rethinking Sustainable Development Goal 7 through struggles over traditional energy practices in Sierra Leone. Energy Policy 105(C): 635-641. |
| [19] |
UNEP 2008. World Database on Protected Areas: Site Information, UN Environment Programme World Conservation Monitoring Centre.
https://www.unep-wcmc.org 14 March, 2025. |
| [20] | Bowden DJ 1997. The geochemistry and development of lateritized footslope benches: The Kasewe Hills, Sierra Leone, Geological Society, London, Special Publications 120(1): 295-305. Pages 295 – 305 |
| [21] | Lytwyna J, Burkea K, Culver S. 2006. The nature and location of the suture zone in the Rokelide orogen, Sierra Leone: Geochemical evidence. Journal of African Earth Sciences 46(5): 439-454. |
| [22] | Pop-Moldovan V, Corcoz L, Stoian V, Moldovan C, Pleșa A, Vaˆ tca˘ S, Stoian V and Vidican R 2022. Models of mycorrhizal colonization patterns and strategies induced by biostimulator treatments in Zea mays roots. Front. Plant Sci. 13: 1–18. Volume 13 - 2022 | |
| [23] | Brundrett, M. C., Piche, Y. & Peterson, L. (1994). A new method for observing the morphology of VAM. Can. J. Bot., 62: 2128-2134. |
| [24] | FAO/LRSP 1972. Technical Report Number 1. Land Use in Sierra Leone: a reconnaissance survey and evaluation for agriculture. Land Resource Survey Project, Freetown, Sierra Leone. |
| [25] | Grandy, A. S., Wieder, W. R., Wickings, K., Kyker-Snowman, E., 2016. Beyond microbes: Are fauna the next frontier in soil biogeochemical models? Soil Biol. Biochem. 102, 40–44. |
| [26] | Jalloh, A., Roy-Macauley, H., Sereme, P., 2011. Major agro-ecosystems of West and Central Africa: Brief description, species richness, management, environmental limitations and concerns. Agriculture, Ecosystems and Environment 157, 5–16. |
| [27] | Sale, V., P. Aguilera, E. Laczko, P. M¨ader, A. Berner, U. Zihlmann, F. Oehl, 2015. Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi, Soil Biol. Biochem. 84: 38–52. |
| [28] | Amara, D. M. K., A Kamara, and Edwin J J Momoh 2013. Soil Fertility Status of Three Chiefdoms in Pujehun District of Southern Sierra Leone. Research Journal of Agricultural Sciences 4(4): 461-464, July-August (2013). DI: 1334-13-2705-2013-119 |
| [29] | Bennett L. T, Mele P, Annett S and Kasel, S. (2010) Examining links between soil management, soil health, and public benefits in agricultural landscapes: An Australian perspective. Agriculture Ecosystems & Environment 139(1): 1-12 |
| [30] | Chalermsri A, Ampornpan L and Purahong W 2020. Seed Rain, Soil Seed Bank, and Seedling Emergence Indicate Limited Potential for Self-Recovery in a Highly Disturbed, Tropical, Mixed Deciduous Forest. Plants 9(1391): 1-14. |
| [31] | Sonon, L. S., Kissel, D. E., and Saha, U 2022. Cation Exchange Capacity and Base Saturation. The University of Georgia College of Agricultural and Environmental Sciences. Circular 1040. |
| [32] | Lukina, N., Tikhonova, E., Danilova, M., Bakhmet, O., Kryshen, A., Tebenkova, D., Kuznetsova, A., Smirnov, A., Braslavskaya, T., Gornov, A., Shashkov, M., Knyazeva, S., Kataev, A., Isaeva, L., and Zukert, N 2019. Associations between forest vegetation and the fertility of soil organic horizons in northwestern Russia, Forest Ecosyst., 6, 1–19, |
| [33] | Aderonke, D. O. and Gbadegesin, G. A. 2013. Spatial Variability in Soil Properties of a Continuously Cultivated Land. African Journal Agricultural Research, 8, 475-483. |
| [34] | Diop I, Ndoye F, Kane A, Krasova-Wade T, Pontiroli A, Do Rego FA, Noba K. and Prin Y: 2015. Arbuscular mycorrhizal fungi (AMF) communities associated with cowpea in two ecological site conditions in Senegal. African Journal of Microbiology Research. 9(21): 1409-1418. |
| [35] | Vieira LC, da Silva DKA, da Silva IR, Gonçalves CM, de Assis DMA, Oehl F. and da Silva GA: 2018. Ecological aspects of arbuscular mycorrhizal fungal communities in different habitat types of a Brazilian mountainous area. Ecological Research. 34: 182–192. 2019 |
| [36] | Samba-Mbaye RT, Anoir CM, Diouf D, Kane A, Diop I, Assigbete K, Tendeng P. and Sylla SN. 2020. Diversity of arbuscular mycorrhizal fungi (AMF) and soils potential infectivity of Vachellia nilotica (L.) P. J. H. Hurter & Mabb. rhizosphere in Senegalese salt-affected soils. African Journal of Biotechnology 19(7): 487-499. |
| [37] | Teixeira, A. F. D. S, Kemmelmeier K, Marascalchi MN, Stürmer SL, Carneiro MAC. and Moreira FM: 2017. Arbuscular mycorrhizal fungal communities in an iron mining area and its surroundings: Inoculum potential, density, and diversity of spores related to soil properties. Ciência e Agrotecnologia 41(5): 511-525. |
| [38] | Sellal Z, Touhami AO, Chliyeh M, Dahmani J, Benkirane R. and Douira A: 2016. Arbuscular Mycorrhizal fungi species associated with rhizosphere of Argania spinosa (L.) Skeels in Morocco. International Journal of Pure Applied Bioscience 4(1): 82-99. |
| [39] | Song J, Chen L, Chen F. and Ye J: 2019. Edaphic and host plant factors are linked to the composition of arbuscular mycorrhizal fungal communities in the root zone of endangered Ulmus chenmoui Cheng in China. Ecology and Evolution 9: 900–8910. |
| [40] | Bini, D.; Santos, C. A.; Carmo, K. B.; Kishino, N.; Andrade, G.; Zangaro, W.; Nogueira, M. A. 2013. Effects of land use on soil organic carbon and microbial processes associated with soil health in southern Brazil. European Journal of Soil Biology 55: 117-123. |
| [41] | Schnoor TK, Lekberg Y, Rosendahl S. and Olsson PA: 2011. Mechanical soil disturbance as a determinant of arbuscular mycorrhizal fungal communities in semi-natural grassland. Mycorrhiza 21: 211-220. |
| [42] | Klironomos JN. and Hart MM: 2002. Colonization of roots by arbuscular mycorrhizal fungi using different sources of inoculum. Mycorrhiza 12: 181- 184. |
| [43] | Giovannetti M, Azzolini D. and Citernesi AS: 1999. Anastomosis Formation and Nuclear and Protoplasmic Exchange in Arbuscular Mycorrhizal Fungi. Applied and Environmental Microbiology 65(12): 5571–5575. |
| [44] | Zhang H, Wei S, Hu W, Xiao L. and Tang M: 2017. Arbuscular Mycorrhizal Fungus Rhizophagus irregularis Increased Potassium Content and Expression of Genes Encoding Potassium Channels in Lycium barbarum. Frontiers in Plant Science 8: 440. |
| [45] | Melo CD, Walker C, Krüger C, Borges PAV, Luna S, Mendonça D, Fonseca HMAC. and Machado AC: 2019. Environmental factors driving arbuscular mycorrhizal fungal communities associated with endemic woody plant Picconia azorica on native forest of Azores. Annals of Microbiology 69: 1309–1327. |
| [46] | Liu J, Liu J, Liu J, Cui M, Huang Y, Tian Y, Chen A. and Xua G: 2019. The potassium transporter SlHAK10 is involved in mycorrhizal potassium uptake. Plant Physiology 180: 465–479. |
| [47] | Nieves-Cordones, M., Alemán, F., Martı́nez, V., and Rubio, F. (2014). K+ uptake in plant roots. the systems involved, their regulation and parallels in other organisms. Journal of Plant Physiol. 171(9), 688–695. |
APA Style
Johnny, J., Lebbie, A., Wadsworth, R. (2025). Soil and Mycorrhizal Diversity and Distribution in Relation to G. copallifera in Kasewe Forest Reserve, Southern Sierra Leone. Plant, 13(3), 124-137. https://doi.org/10.11648/j.plant.20251303.12
ACS Style
Johnny, J.; Lebbie, A.; Wadsworth, R. Soil and Mycorrhizal Diversity and Distribution in Relation to G. copallifera in Kasewe Forest Reserve, Southern Sierra Leone. Plant. 2025, 13(3), 124-137. doi: 10.11648/j.plant.20251303.12
@article{10.11648/j.plant.20251303.12,
author = {Jonathan Johnny and Aiah Lebbie and Richard Wadsworth},
title = {Soil and Mycorrhizal Diversity and Distribution in Relation to G. copallifera in Kasewe Forest Reserve, Southern Sierra Leone
},
journal = {Plant},
volume = {13},
number = {3},
pages = {124-137},
doi = {10.11648/j.plant.20251303.12},
url = {https://doi.org/10.11648/j.plant.20251303.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.plant.20251303.12},
abstract = {Gibourtia copallifera is a rare, range-restricted tree native to the Kasewe Forest Reserve in southern Sierra Leone. Historically exploited for gum copal, it now faces threats from charcoal production due to its high-quality charcoal. Although the species shows good growth in forest patches established 60 years ago, natural regeneration remains minimal. While many tropical tree species form beneficial relationships with arbuscular mycorrhizal fungi (AMF), these associations are still poorly understood in humid regions like Sierra Leone. This study investigated the AMF associations of G. copallifera and assessed changes in mycorrhizal diversity in relation to forest degradation and tree development stages. Soil characterization was conducted through profile analysis, and samples were subjected to laboratory testing. Three soil types were identified within the reserve: K 01—barren land with sparse vegetation; K 02—forest land dominated by G. copallifera; and K 03—upland fallow agricultural land with shrubs and a few trees. Soil analysis indicates pH values ranging from 5.03 to 5.87 (acidic), with calcium as the most dominant exchangeable base, followed by magnesium, potassium, and sodium. The surface horizon under G. copallifera exhibited the highest total exchangeable bases, and high cation exchange capacity (CEC) was linked to the presence of decomposed plant matter. A total of 22 AMF species were identified in plant root samples, with a significantly higher proportion found in non-degraded forest patches. These accounted for around 68% of AMF species, especially Scutellospora and Gigaspora, along with Glomus and Acaulospora. AMF species richness and diversity were considerably higher in non-degraded patches (3.13 species) than in degraded areas (1.75). Mycorrhizal frequency and intensity were also significantly greater in undisturbed forest sites. AMF colonization peaked in mature trees and was lowest in seedlings, although the variation across trials was not statistically significant (P = 0.07). The study concludes that K01 is unsuitable for G. copallifera due to its shallow depth, which restricts growth. In contrast, K02 and K03 are more favorable for the species. However, many K02 areas have been converted to agricultural land, and in locations where natural forest regeneration is allowed, more aggressive species such as Gmelina arborea and Anisophyllea laurina tend to dominate. Additionally, AMF play a crucial role in promoting the growth of G. copallifera in nutrient-poor soils. These findings are important for informing reintroduction and reforestation strategies for this native tropical tree species.
},
year = {2025}
}
TY - JOUR T1 - Soil and Mycorrhizal Diversity and Distribution in Relation to G. copallifera in Kasewe Forest Reserve, Southern Sierra Leone AU - Jonathan Johnny AU - Aiah Lebbie AU - Richard Wadsworth Y1 - 2025/07/09 PY - 2025 N1 - https://doi.org/10.11648/j.plant.20251303.12 DO - 10.11648/j.plant.20251303.12 T2 - Plant JF - Plant JO - Plant SP - 124 EP - 137 PB - Science Publishing Group SN - 2331-0677 UR - https://doi.org/10.11648/j.plant.20251303.12 AB - Gibourtia copallifera is a rare, range-restricted tree native to the Kasewe Forest Reserve in southern Sierra Leone. Historically exploited for gum copal, it now faces threats from charcoal production due to its high-quality charcoal. Although the species shows good growth in forest patches established 60 years ago, natural regeneration remains minimal. While many tropical tree species form beneficial relationships with arbuscular mycorrhizal fungi (AMF), these associations are still poorly understood in humid regions like Sierra Leone. This study investigated the AMF associations of G. copallifera and assessed changes in mycorrhizal diversity in relation to forest degradation and tree development stages. Soil characterization was conducted through profile analysis, and samples were subjected to laboratory testing. Three soil types were identified within the reserve: K 01—barren land with sparse vegetation; K 02—forest land dominated by G. copallifera; and K 03—upland fallow agricultural land with shrubs and a few trees. Soil analysis indicates pH values ranging from 5.03 to 5.87 (acidic), with calcium as the most dominant exchangeable base, followed by magnesium, potassium, and sodium. The surface horizon under G. copallifera exhibited the highest total exchangeable bases, and high cation exchange capacity (CEC) was linked to the presence of decomposed plant matter. A total of 22 AMF species were identified in plant root samples, with a significantly higher proportion found in non-degraded forest patches. These accounted for around 68% of AMF species, especially Scutellospora and Gigaspora, along with Glomus and Acaulospora. AMF species richness and diversity were considerably higher in non-degraded patches (3.13 species) than in degraded areas (1.75). Mycorrhizal frequency and intensity were also significantly greater in undisturbed forest sites. AMF colonization peaked in mature trees and was lowest in seedlings, although the variation across trials was not statistically significant (P = 0.07). The study concludes that K01 is unsuitable for G. copallifera due to its shallow depth, which restricts growth. In contrast, K02 and K03 are more favorable for the species. However, many K02 areas have been converted to agricultural land, and in locations where natural forest regeneration is allowed, more aggressive species such as Gmelina arborea and Anisophyllea laurina tend to dominate. Additionally, AMF play a crucial role in promoting the growth of G. copallifera in nutrient-poor soils. These findings are important for informing reintroduction and reforestation strategies for this native tropical tree species. VL - 13 IS - 3 ER -
Department of Wildlife Management and Conservation, Njala University, Njala, Sierra Leone
Department of Biological Sciences, Njala University, Njala, Sierra Leone, West Africa Plant List Authority, IUCN Species Survival Commission, Gland, Switzerland
Department of Biological Sciences, Njala University, Njala, Sierra Leone
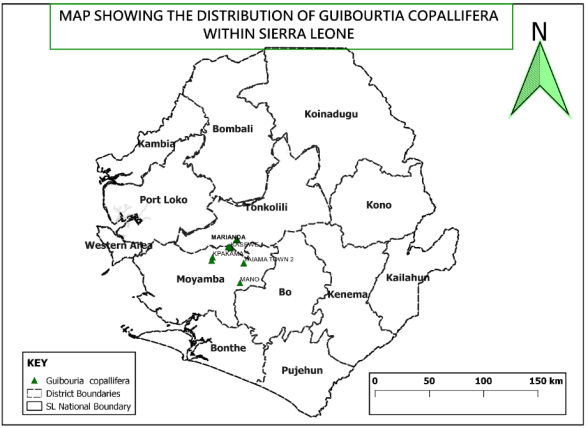
Figure 1. Map of Sierra Leone showing distribution of the species in the districts.

Figure 2. Pictures of the soil profiles.
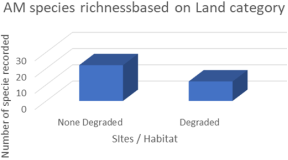
Figure 3. AM species richness based on land quality.
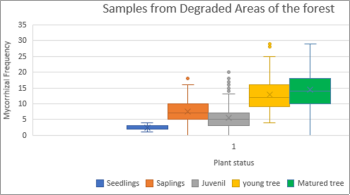
Figure 4. Box-plots showing frequency of colonization within growth stages of the plants (degraded sites).
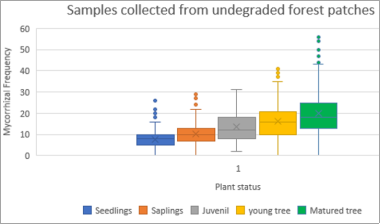
Figure 5. Box-plots of root colonization with growth form (undisturbed sites).
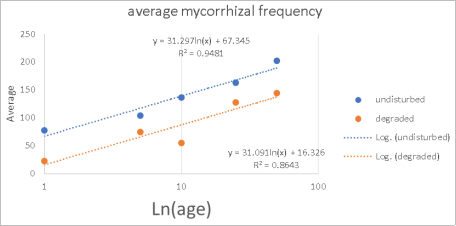
Figure 6. Comparison of colonization rates with approximate growth stage of plants.



